|
| |||||||||||||||||
|
ANATOMIE VAN HET ADEMHALINGSSTELSEL BIJ DE CAVIA Het ademhalingsstelsel van de cavia heeft veel diersoort-specifieke kenmerken. Deze worden uitgelegd met behulp van gesimplificeerde tekeningen. Verder bespreken wij de hoofdzaken van het functioneren (fysiologie) van het ademhalingsstelsel. Voor meer details verwijzen wij naar de gespecialiseerde naslagwerken.
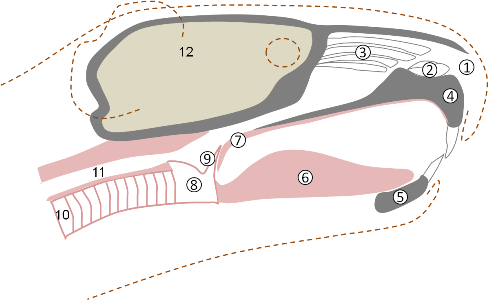
De nasofarynx is het bovenste gedeelte van de luchtwegen. Het is de ruimte die zich boven het gehemelte bevindt. Het omvat de neusholte en reikt tot aan het strottenhoofd (larynx). Een versimpeld schema van de nasofarynx van de cavia is weergegeven in Fig. 1. De volgende structuren zijn aangegeven: Naris. (1) Dit is de uitwendige neusopening. Neusschelpen (conchae nasales). (2-3) Dit zijn (kraak)benige structuren die de neusholte in verschillende compartimenten - neusgangen - verdelen. In de onderste neusgang (meatus nasi ventralis) mondt de traanbuis uit. De traanbuis voert de overtollige traanvocht af naar de neusholte. Verstopping van de traanbuis treedt op bij infecties van de bovenste luchtwegen; door verminderde afvoer van traanvocht ontstaat er traanoog. De neusschelpen hebben een groot oppervlak en zijn bedekt met reukepitheel. Het reukepitheel is sterk geïnnerveerd; zenuwbundels (filae olfactoriae) transporteren de informatie doorheen de lamina cribriformis (het poreuze deel van de aansluitende botplaat - os ethmoidale) naar de hersenen. Reuk is zeer goed ontwikkeld bij cavia's. De structuren in de neusholte zijn sterk doorbloed en spelen een rol in warmteuitwisseling. Reukepitheel bevat trilharen. De beweeglijke trilharen houden stofdeeltjes, bacteriën en andere schadelijke partikels tegen. Op deze manier komt er niet zo veel vervuiling in de longen. Bijholten (sinus paranasales) zijn praktisch afwezig. Sinusitis (een hardnekkige infectie, en een sequeel van bovenste-luchtweginfecties) is dus geen probleem bij cavia's. Boven- en onderkaak, tong. (4-6) Deze zijn aangegeven ter oriëntatie. Zacht gehemelte (palatinum molle). (7) Dit is de verlenging van het benige harde gehemelte. Bij cavia's is het zachte gehemelte opvallend lang; zacht gehemelte en epiglottis (9) scheiden de mondholte vrijwel volledig af van de nasofarynx. Tubae auditivae. In de zijwanden van het achterste gedeelte van de nasofarynx monden de inwendige gehoorgangen uit (tubae auditivae, niet getekend). Deze zijn klinisch zeer relevant (middenoorontsteking als een sequeel van bovenste-luchtweginfecties). Larynx en epiglottis. (8-9) Het strottenhoofd (larynx) is een kraakbenige buis die de nasofarynx met de luchtpijp verbindt. In de larynx bevinden zich de stembanden (niet getekend). De laryngeale opening bevindt zich net achter het strotklepje (epiglottis) (9). Bij het slikken buigt de epiglottis zich naar achter, waardoor hij de laryngeale opening afsluit. Dit voorkomt dat voedsel in de luchtpijp terechtkomt. Bij cavia's is er veel overlap tussen de epiglottis en het zachte gehemelte (zoals bij paarden). Dit specifieke anatomische kenmerk heeft talrijke consequenties:
Luchtpijp (trachea) en slokdarm. (10-11) Deze zijn gegeven ter oriëntatie. Slokdarm bevindt zich dorsaal van (achter) de luchtpijp in de nekregio; in de borstholte gaat het zich steeds meer naar links verplaatsen.
De infecties ter hoogte van de nasofarynx noemen wij de infecties van de bovenste luchtwegen: URI of the upper respiratory infections (Bovenste luchtwegen). Het is een zeer bekend ziektebeeld bij cavia's. Er bestaat een genetische predispositie voor URI en overige luchtweginfecties. Bij cavia's met een korte neus (brachycefale cavia's) is de neuspassage relatief kort (Fig. 2). Deze dieren zijn dus niet zo goed in staat de lucht van de stofdeeltjes en microben te zuiveren. Chronische en/of terugkerende luchtweginfecties zien wij voornamelijk bij cavia's met een korte neus. De infecties verlopen meestal veel acuter, en dodelijke complicaties (longontsteking en -abcessen) komen veel vaker voor dan bij overige cavia's. Helaas wordt de korte neus door vele fokkers als een zeer gewild raskenmerk beschouwd, zodat er veel brachycefale cavia's gekweekt worden.
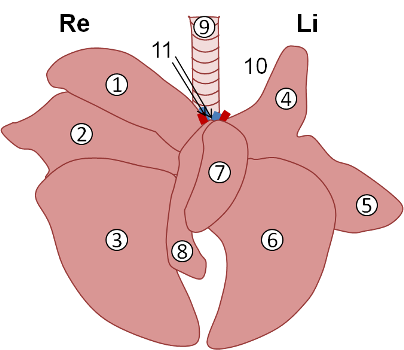
De structuur. Een overzicht van de onderste luchtwegen is gegeven in Fig. 3. De lucht komt binnen via de luchtpijp, die zich splitst bij de ingang naar de long (de longhilus) in twee principale bronchiën - de linke en de rechte. Daarna vertakken de principale bronchiën zich verder in steeds kleinere buisjes: lobaire bronchiën (één voor iedere longkwab; de long van de cavia bestaat uit 7 kwabben, zoals afgebeeld in Fig. 3), segmentale bronchiën en bronchioli. De grotere luchtwegen - trachea t/m segmentale bronchiën - zijn verstevigd door kraakbeen (zie de kraakbeenringen van de trachea in Fig. 1 en 3). In de bronchiale wand bevindt zich het lymfoïde weefsel (BALT, of bronchus associated lymphoid tissue). Dit is belangrijk voor het afweren van de infecties die via de luchtwegen intreden. Bronchiën en bronchioli bevatten glad spierweefsel, dat onder controle van het autonome zenuwstelsel staat. Daardoor is verwijding en vernauwing van de luchtwegen mogelijk. De binnenwanden van de buisjes zijn bekleed met het respiratoir epitheel met trilharen en slijmbekercellen. Door de continue beweging van de trilharen en door de uitscheiding van slijm worden stof en andere partikels actief weggewerkt. Het kleinste buisje - bronchiolus respiratorius - staat in contact (via ductus alveolaris) met de longblaasjes (alveoli). Een histologische snede door het longweefsel is afgebeeld in Fig. 4. De wand van longblaasjes is zeer dun en bestaat uit afgeplatte epitheelcellen (pneumocyten type I), die in direct contact staan met de wand van de capillaire bloedvaten. Op deze manier kan er gasuitwisseling (opname van zuurstof en uitscheiding van koolstof dioxide) plaatsvinden. Longblaasjes bevatten ook steuncellen (pneumocyte type II) die een oppervlakte-actieve stof - surfactant aanmaken. Overige cellen zijn macrofagen die de cel- en bacterieresten opruimen, en gespecialiseerde sensorische en endocrine cellen. De longfunctie. De longfunctie hangt af van de efficiëntie van het opvullen van de longblaasjes met lucht (ventilatie) en van het transporteren van zuurstof door het bloed. Het opvullen van de blaasjes is mogelijk dankzij de aanwezigheid van het surfactant. Het surfactant verlaagt de oppervlaktespanning van de fysiologische vloeistof, waardoor de lucht-vloeistof grenslaag blijft bestaan. Zonder surfactant zouden de blaasjes collapsen en verkleven. Alle chemische middelen die een anti-surfactant werking hebben (simeticon, dimeticon) zijn dus levensgevaarlijk als ze per ongeluk in de longen terechtkomen. Voor een goede vulling moet het longweefsel soepel zijn, met andere woorden, de long moet een hoge compliantie hebben. Veel bindweefsel afzetting (chronische ontstekingen, oude letsels) vermindert de compliantie en benadeelt de longfunctie. Voor een goede gasuitwisseling moet de lucht-bloed barrière zo dun mogelijk blijven. De aanwezigheid van vloeistof in de longblaasjes (longoedeem - bij longontsteking of bij hartinsufficiëntie) maakt de barrière dikker en verslechtert de gasuitwisseling. Verdikking van de alveolaire wanden (bij ontsteking) veroorzaakt dezelfde problemen. Voor de efficiënte zuurstofopname moet de doorbloeding (perfusie) van de long goed zijn. Slechte perfusie (onvoldoende bloedstroming) zoals bij hartfalen of in shocktoestanden, leiden tot zuurstofnood. De specifieke ziektebeelden worden verder uitgewerkt in Onderste luchtwegen.
DE BORSTHOLTE ALS GEHEEL De borstholte (thorax) bevat de longen, het hart, de leidingen (slokdarm, bloedvaten, zenuwen) en veel vliezige steunstructuren. Een versimpelde schematische weergave van de borstholte vindt u in Fig. 5. De borstholte is begrensd door de ribbenkast (1) en het middenrif (diaphragma) (2). De borstholte kan (grof) verdeeld worden in drie compartimenten: de pleuraholte (4), het hartzakje (pericardium) en de overige ruimte (mediastinum) (5). De pleuraholte (4) wordt gevormd door twee bladen borstvlies (pleura): de buitenste - pariëtale pleura (6) - die vastzit aan de wand van de borstkas, en de binnenste - viscerale pleura (7) - die de longen bekleedt. De holte bestaat uit twee delen - linker en rechter - die niet in verbinding met elkaar staan. De pleuraholte is een virtuele ruimte (op de tekening is het overdreven groot geschetst); dit wil zeggen dat er in de normale toestand de twee borstvliesbladen nagenoeg tegen elkaar aan liggen. Er is een kleine vrije ruimte tegen het middenrif en tegen het mediastinum - de zogenaamde recessus costodiaphragmaticus en recessus costomediastinalis. Recessi dienen als een reserveruimte, zodat de longen zich meer dan normaal kunnen opvullen bij verhoogde behoefte aan ventilatie. Bij een gezond dier is er slechts een kleine hoeveelheid vocht in de pleuraruimte. De hoeveelheid vocht neemt sterk toe in ziekte; zo ontstaat er hydrothorax. De vocht kan helder en eiwitarm zijn (transsudaat) bij hartfalen, extreme uithongering, lever- en nierfalen, of troebel en eiwitrijk (exsudaat) bij ontstekingsprocessen. Bij ernstige longaantasting door bacteriën kan er ook etter, bloed en zelfs lucht in de pleuraholte komen. Uiteraard kunnen de longen zich niet meer opvullen en uitzetten, waardoor er acute ademnood ontstaat. Lees verder in Onderste luchtwegen. Het hartzakje (pericardium) bevat het hart (9). Het hart ligt ventraal van de long, maar wordt deels omhuld door de longkwabben die ventraal buigen. In dit opzicht is de tekening in Fig. 5 niet realistisch: het is een platte projectie, terwijl de structuren driedimensionaal zijn. Het hart ligt nagenoeg los in de borstholte; het wordt enkel opgehangen aan de grote bloedvaten: aorta (11), truncus pulmonalis en zijn longarteriën (12), vena cava (10) en de longvenen. Er kan eventueel een zwak aangelegd ligament aanwezig zijn, dat het hartzakje met het middenrif verbindt. De structuur en functie van het hart wordt beschreven in andere hoofdstukken (Circulatie). Het mediastinum (5) is de overgebleven ruimte. Hier bevinden zich de grote leidingen: de slokdarm, de grootste slagader (aorta) en ader (v. cava caudalis) van het lichaam, de grootste lymfevaat (ductus thoracicus, in de buurt van de aorta), de mediastinale lymfeknopen en nervus phrenicus (zie verder). Mediastinum bevat een kleine hoeveelheid extracellulaire vloeistof. De aanwezigheid van veel vocht, etter, bloed of lucht (door ziekte of trauma) leidt tot ernstige ademhalingsproblemen.
Het aanzuigen van lucht is enkel mogelijk als de druk in de borstholte lager is dan de buitendruk (1 atmosfeer). De druk in de borstholte wordt verlaagd door de contracties van de ademhalingsspieren. De belangrijkste ademhalingsspier is het middenrif (diaphragma) - de scheiding tussen de borst- en de buikholte. Het ontspannen middenrif puilt uit in de borstholte (Fig. 5); door de contractie wordt het platter, waardoor de borstholte vergroot. Door volumevergroting ontstaat er onderdruk en de lucht wordt aangezogen. Bij een sterk vergrote maag (gas, tympanie) duwt de maag het middenrif in de richting van de borstholte, waardoor het aanzuigen van lucht moeilijk wordt. Gasophoping in de maag en benauwdheid zien wij dus vrijwel altijd samen. Meestal is het een vicieuze cirkel: bij ademnood wordt er veel lucht ingeslikt en dit leidt tot verdere verslechtering van de ademhaling. Overige spieren die aan de vergroting van de borstholte bijdragen zijn: mm. intercostales externi (korte spieren die de ribben met elkaar verbinden aan de buitenkant van de ribbenkast), mm. levatores costarum (zeer korte spieren die de wervels met de ribben verbinden) en m. rectus thoracis (diepe borstspierlaag). Het uitademen van lucht gebeurt hoofdzakelijk automatisch (door relaxatie van de bovengenoemde spieren), maar het kan ook ondersteund worden door de samentrekking van mm. intercostales interni (analoog aan intercostales externi, maar aan de binnenkant van de ribbenkast) en m. transversus thoracis. Bij een geforceerde uitademing gebruikt het dier ook de buik- en lendespieren (m. obliquus abdominis externus, m. latissimus dorsi, m. psoas major en andere). Bij ernstige zuurstofnood persen de cavia's lucht uit de longen om meer lucht te kunnen aanzuigen en om de stapeling van koolstof dioxide tegen te gaan. Men ziet dan heftige samentrekkingen van de buikspieren, waardoor de achterhand van de cavia duidelijk meebeweegt tijdens het uitademen. Dit noemen we "de buikspierpomp". Het geforceerd ademen gebeurt uiteraard enkel in ziekte: bij long- en circulatieproblemen en bij metabole acidose (bloedverzuring door abnormale stofwisselingsprocessen, bv in zwangerschapsvergiftiging). Het basisritme van de ademhaling wordt onwillekeurig opgelegd door het pneumotaxisch centrum in de hersenstam (pons). Dit centrum controleert het ademhalingscentrum. Het ademhalingscentrum bestaat uit het inspiratie centrum (de dorsale groep) en het expiratie centrum (de ventrale groep) in het verlengde merg, of preciezer in het formatio reticularis. Formatio reticularis is en netwerk van zenuwcellen, betrokken bij het bewustzijn, pijnsensatie, controle van hartwerking en andere functies. Bij de normale (niet geforceerde) ademhaling wordt enkel de dorsale groep ingeschakeld. Vanuit de dorsale groep vertrekken de signalen periodiek naar de cellichamen van nervus phrenicus (de belangrijkste) en nervi intercostales. Nervus phrenicus verlaat het ruggenmerg ter hoogte van C3 (de derde halswervel) en loopt door de borstholte naar het middenrif toe; de linker zenuw gaat langs het linkerhart, rechter zenuw verloopt samen met v. cava caudalis. N. phrenicus stuurt het middenrif aan. Door de samentrekking van het middenrif en de overige ademhalingsspieren (gestuurd door nn. intercostales) wordt het inademen geïnitieerd. Overmatige stimulatie van n. phrenicus resulteert in hik; dit ziet men vaak bij cavia's (is een onschuldig verschijnsel). Het uitademen gebeurt in normale omstandigheden passief en treedt op als het zenuwsignaal uit het inspiratie centrum stopt. Enkel bij een verhoogde behoefte aan ventilatie wordt de ventrale groep (het expiratie centrum) ook gestimuleerd, waardoor de lucht actief uit de longen gepompt wordt. De ventrale groep, met name de groep neuronen die de zogenaamde pre-Bötzinger complex vormen, heeft een overal stimulerend effect zowel op het inademen als op het uitademen. De klinische relevantie hiervan is dat bepaalde factoren, die de werking van dit gedeelte van de hersenen beïnvloeden, tot ademhalingsdepressie en verstikkingsdood kunnen leiden. Een bekend voorbeeld is ademhalingsdepressie in de narcose toestand. Alle bekende anaesthesiemiddelen veroorzaken depressie van het ademhalingscentrum. Dit is een onvermijdelijke bijwerking, omdat de anaesthesiemiddelen de zenuwcellen in het formatio reticularis beïnvloeden. Formatio reticularis is immers verantwoordelijk voor (onder andere) het doorgeven van pijnsignalen naar het bewuste deel van de hersenen; het uitschakelen van deze verbindingen is het werkprincipe van anaesthesiemiddelen. Andere bekende oorzaken van ademhalingsdepressie zijn: hersentrauma en schade aan het ruggenmerg in de halsregio (na valpartijen). Hersentrauma leidt tot de zwelling van het hersenweefsel, waardoor het ademhalingscentrum niet meer kan functioneren. Schade aan het ruggenmerg hoog in de halsregio (trauma) veroorzaken de verlamming van n. phrenicus. Dergelijke schade is meestal dodelijk. Het basispatroon van de ademhaling wordt beïnvloed door vele factoren:
REFERENTIES Adriaensen D, Scheuermann DW, 1993, Neuroendocrine cells and nerves of the lung, The Anatomical Record 236: 70-85. Guyton AC, Hall JE, Textbook of Medical Physiology, Xth edition, Saunders publishing 2000. Popesko P, Rajtova V, Horak J, A Colour Atlas of Anatomy of Small Laboratory Animals, vol. 1 Rabbit, Guinea Pig, Saunders publishing 2002. Salinger S, 1948, The Paranasal Sinuses, Arch. Otolaryngol. 48: 430-462. | |||||||||||||||||
| | |||||||||||||||||